DNA replication involves the copying of each strand of the double helix to give a pair of daughter strands. Replication begins at a specific sequence, called the origin. After initiation begins at an origin sequence, all sequences are replicated no matter what their information. This principle leads to the idea of molecular cloning, or recombinant DNA. Cloning enables the production of a single DNA sequence in large quantities.
A recombinant DNA consists of two parts: a vector and the passenger sequences. Vectors supply replication functions—the origin sequences to the recombinant DNA molecule. After it joins to a vector, any passenger sequence can be replicated. The process of joining the vector and passenger DNAs is called ligation. DNA ligasecarries out ligation by using ATP energy to make the phosphodiester bond between the vector and passenger. If the vector and passenger DNA fragments are produced by the action of the same restriction endonuclease, they will join by base-pairing. After they are ligated to a vector, it is possible to make an essentially unlimited amount of the passenger sequence.
Plasmid vectors
Plasmids are circular DNAs that are capable of independent replication. Many naturally occurring bacteria contain plasmids; plasmid vectors are derived from naturally occurring plasmids. Plasmid vectors have several properties. First, they contain single restriction sites for several enzymes. Cleaving with one of these enzymes generates a single, linear form of the plasmid. This feature helps to ensure efficient ligation, because every ligation product will contain the entire vector sequence. Secondly, vectors are made to contain selectable genetic markers so that cells which contain the vector can be propagated. For example, many plasmid-cloning vectors contain a gene that encodes resistance to the antibiotic ampicillin (a member of the penicillin family). Cells that contain the vector can then be selected for by growth in the presence of the antibiotic; cells that don't contain the vector sequence will be inhibited by ampicillin in the growth medium, as shown in Figure 1 .
| |||
| |||
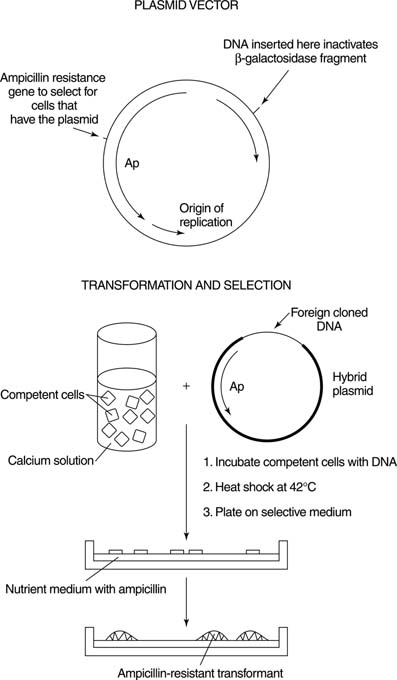
Thirdly, a means to detect which cells have only the plasmid vector as opposed to the recombinant product must exist. This determination is usually accomplished by a mechanism called insertional inactivation. The idea of insertional inactivation is that inserting passenger DNA into a gene interrupts the sequence of the gene, thereby inactivating it. For example, the restriction sites of many common plasmid-cloning vectors are located in a fragment of a gene for β-galactosidase, an enzyme involved in lactose metabolism. When cells containing just the vector are grown in the presence of an artificial substrate related to lactose, the colonies turn blue, because active enzyme is made. On the other hand, when the restriction site has a piece of foreign DNA inserted into it, the gene cannot make an active protein fragment because the DNA sequence interrupts the coding sequence of the gene. As a result, colonies of the bacteria that contain cloned foreign DNA appear whitish. The bacteria that are present in the colony can be grown separately, and standard biochemical procedures easily isolate the recombinant DNA they contain. If no such selection existed, each colony would have to be grown separately and its DNA analyzed—a very “hit or miss” proposition in many cases.
After a recombinant plasmid has been formed in the laboratory, it must be replicated. This process begins with the growth of the cell containing the recombinant plasmid. Plasmids are usually transferred to new hosts bytransformation. Transformation is the addition of naked foreign DNA into a recipient bacterium. Because the plasmid usually contains a gene encoding the ability to grow in the presence of an antibiotic, identifying a recombinant bacterium is a two-part process. First, growth in the presence of the antibiotic ensures that each bacterial colony contains a plasmid, while the color of the colony identifies the plasmids that contain inserted DNA. Finally, the growth of the transformed bacterial colony amplifies the clones, that is, makes more copies of the plasmid. These steps can be done with a known piece of DNA or with a less-defined collection of insert DNA— for example, all the DNA from an organism. A collection of different cloned DNAs is called a library. The number of independent sequences in a library is called its complexity; the more complex the library, the greater number of independent sequences it contains.
Other types of cloning vectors
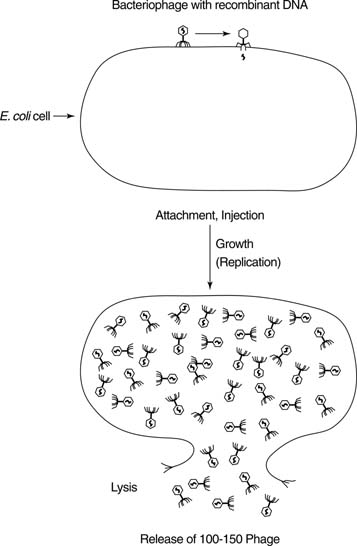
Viruses that infect bacteria are called bacteriophages. Native bacteriophage have been formed into vectors that can also be used for cloning. Bacteriophage vectors have three advantages over plasmids. First, they can carry significantly more foreign DNA than can plasmids, which are limited to about 5,000 base pairs (5 kilobases) of foreign DNA. Plasmids with larger-sized inserts tend to be unstable when amplified. The larger the insert, the fewer independent clones are required to have a reasonable chance of identifying any single gene or sequence in the collection of cloned DNA. Secondly, the virus particles of bacteriophage vectors can accept DNA only of a narrow size range. This means that DNA can be preselected so that each recombinant virus will contain only a single foreign sequence. This property is a consequence of the fact that DNA (whether native or foreign) must be packaged into a protein coat to be infectious. DNAs that are too large or too small can't be packaged efficiently, so they will not be represented in the library. Finally, bacteriophage infection can be a very efficient process, with nearly 100 percent of the packaged virus particles leading to a productive infection. In contrast, cells take up only one of 100,000 plasmid molecules in a transformation procedure. Bacteriophage clones are amplified by repeated cycles of infection and growth. See Figure 2 .
| |||
|
0 comments:
Post a Comment