The biochemistry of the lactose ( lac) operon explains many principles of regulation. The lac operon encodes a set of genes that are involved in the metabolism of a simple sugar, lactose. Lactose is a disaccharide composed of two sugars (galactose and glucose) with a β-linkage between carbon 1 of galactose and carbon 4 of glucose, as shown in Figure 1 .
| |||
| |||
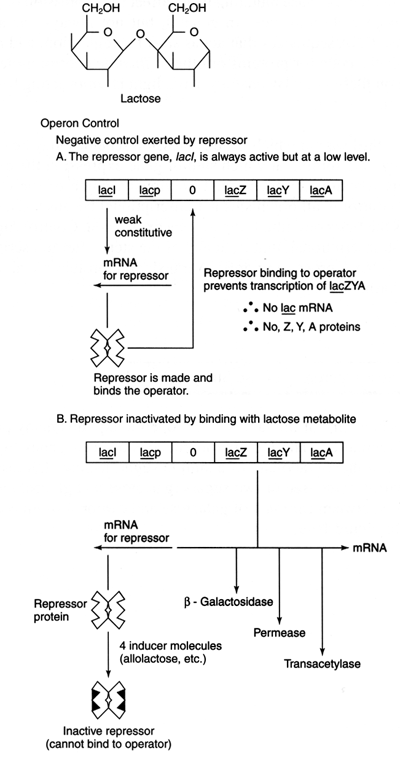
The lac operon encodes three proteins: β-galactosidase (the product of the lacZgene), lactose permease (the product of the lacY gene), and lactose transacetylase (the product of the lacA gene). The function of lacA is not known, but a mutation in either lacZ or lacY means that the cell can't grow by using lactose as a sole carbon source. All three structural genes are transcribed from a common promoter site, in the direction Z-Y-A. The lac transcript is termed polycistronic because it contains more than one coding sequence.
Closely linked to the lac structural genes is the gene ( lacI) for the lac repressor, a tetramer of four identical subunits. The repressor has two functions. First, it binds to the DNA near the lac promoter and prevents transcription of the structural genes. Secondly, it binds to a small molecule called an inducer. In the cell, the inducer is allolactose, a metabolite of lactose. The binding of inducer to the repressor is cooperative, meaning that the binding of one molecule of inducer makes binding of the next one more favorable, and so on. This means that the repressor binds inducer in an all-or-none fashion.
In the absence of inducer, the repressor protein binds to a sequence called theoperator ( lacO) which partially overlaps with the promoter. When it is bound to the operator, lac repressor allows RNA polymerase to bind the promoter and form an open complex but not to elongate transcription. Repressor is therefore a negative regulator of gene expression: If repressor is not present (for example, the bacterium is deleted for the lacI gene), then transcription of the lac genes occurs, and the structural genes are expressed whether or not inducer is present. The unregulated, “always on” expression caused by a lacI- mutation is calledconstitutive expression. This behavior is characteristic of a negative control element. LacO is also a negative element. Deletion of lacO leads to constitutive expression of the lac genes. The difference between the two types of constitutive mutation is seen when the genes are put into an artificial situation where two copies of the relevant control genes exist. See Figure 2 .
| |||
| |||

If a cell is provided with one wild-type and one mutant lacI gene, then the “good” (wild-type) copy of the repressor gene will provide functioning repressor to the other operator, and expression will be normally controlled. This behavior is characteristic of a diffusible control element, and the repressor is said to act intrans. If the same experiment is carried out with one wild-type and one mutant lacO gene, then the genes controlled by the mutant lacO will be constitutively expressed and the genes controlled by the wild-type copy of lacO will be normally regulated. This is characteristic of a site where the product of another gene acts, and lacO is therefore termed cis-acting.
This model also accounts for the behavior of mutants that cause repressor to fail to bind inducer. If the repressor can't bind inducer, then the lac genes it controls are permanently turned off because repressor will be bound to the operator whether inducer is present or not. In a diploid situation, both sets of genes will be turned off, because the repressor will bind to both operators.
A second level of control is superimposed on the repressor-operator interaction described previously. In vivo, lac gene expression is greatly reduced by the presence of glucose in the medium, even if enough lactose is present to release the repressor from the operator. This makes good metabolic sense. Glucose is more easily catabolized (broken down) than is lactose, which must be broken down into glucose and galactose, followed by the specialized pathway for galactose metabolism. The phenomenon by which glucose reduces the expression of the lacoperon is called catabolite repression, as shown in Figure 3 .
| |||
| |||

Catabolite repression is a two-part system. The first component is the small-molecule regulator, cyclic AMP. Glucose decreases cyclic AMP synthesis. The second component is cyclic AMP binding protein, CAP. CAP binds cAMP and thereby helps RNA polymerase bind to the promoter. When bound to cAMP, CAP binds to a sequence at the 5′ end of the lac promoter. CAP binding bends the DNA, allowing protein-protein contact between CAP and polymerase. It therefore behaves in the opposite manner of repressor. Repressor (LacI) binds to operator DNA only in the absence of its small-molecule ligand, while CAP binds to promoter DNA in the presence of its small-molecule ligand.
These two complementary systems allow the bacterial cell to metabolize lactose in response to two stimuli. “Switching on” the expression of the lac operon requires both the absence of glucose and the presence of lactose. This series of switches allows complex expression patterns to be built up from simple components. For this reason, the lac system is a model for other, apparently more complex, biological control systems, such as hormone action or embryonic development.
0 comments:
Post a Comment