DNA information is subject to loss or degradation. After all, DNA is a chemical, and chemicals can undergo reactions. Damage to a DNA leads to a loss or alteration of its information content, a process known as mutation. Compounds or agents that cause mutations are called mutagens.
Several types of mutations exist, depending on the changes to the sequence information of the chromosome. Transition mutations are defined as the conversion of G-C to A-T or A-T to G-C base pairs. For example, the compound hydroxylamine causes the loss of the amino group of deoxycytosine in DNA, converting it to deoxyuracil. When replicated, deoxyuracil is a template for deoxy-A incorporation. Hydroxylamine is a monodirectional mutagen, only converting G-C → A-T base pairs.
Transversion mutations convert purines to pyrimidines or vice-versa. For example, conversion of an A-T base pair to a T-A or C-G base pair is a transversion. Ultraviolet light can cause transversions, although not exclusively. Frameshiftmutations result from the insertion or deletion of a single base pair. These are often due to planar intercalating mutagens, which insert into the double helix. More extensive deletions and insertions are common events, as shown in Figure 1 .
| |||
| |||

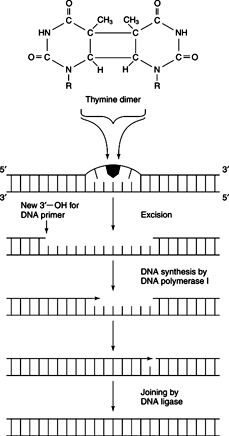
Another important mutagen is ultraviolet light. Recent concern about the depletion of the atmospheric ozone layer by chlorofluorocarbon compounds (CFCs) is due to the role of the ozone in absorbing UV radiation before it can cause mutations in the organisms at the earth's surface. All the DNA bases efficiently absorb UV and become chemically reactive as a result. The formation of pyrimidine dimers from adjacent thymidine residues in DNA interferes with replication and transcription of DNA. See Figure 2 .
| |||
| |||
Mutations can also be “spontaneous,” due to errors in replication, background chemical reaction, and so forth. Because the rates of most chemical reactions increase with increasing temperature, and because mutagenesis is a chemical reaction, not surprisingly, even heat can be thought of as a mutagen.
DNA repair systems
Repair systems in the cell counteract mutations. If no way existed to prevent or revert mutations, life would be too unstable for one generation to give rise to another. Usually, the spontaneous mutation rate of an organism is less than one mutation per billion or so nucleotides per generation. Because humans have about three billion bases of DNA, even identical twins differ by several mutations. Viruses usually don't have DNA repair systems; they keep going from one generation to another either by inserting into a host's DNA or by releasing so many viruses per generation that enough good copies survive to select the active ones at the next generation.
A large number of systems counter mutagenic events. Direct reversal of the mutagenic event is possible in some cases. For example, methylation of guanine by certain DNA alkylating compounds leads to mutation. This form of DNA damage is counteracted by the synthesis of specific proteins that accept the methyl group from the damaged DNA and then are themselves degraded.
Ultraviolet light causes photodimerization of adjacent pyrimidine residues in DNA, such as the formation of thymidine-thymidine dimers.
Photoreaction can remove thymidine-thymidine photodimers. Visible light activates the enzyme DNA photolyase. This enzyme absorbs visible light and transfers that energy to the photodimer, reversing the dimerization directly.
Excision repair is a more general mode of counteracting mutagenic events. Many DNA alterations, including photodimerization, are reversed by cutting out the altered base and its neighbors, followed by synthesis of new DNA by DNA polymerase. In these processes, the double helix provides the information pointing to the “correct” DNA sequence. The initial event is the recognition of the altered DNA segment. For example, formation of pyrimidine dimers changes the shape of the double helix. Repair systems recognize the altered shape to start the repair process. After the repair systems detect the change, nuclease action near, but not necassarily at, the site of mutation cleaves the damaged chain. The cleaved segment is unwound extended, leaving a single-stranded gap. Then DNA polymerase I fills in the gap and DNA ligase closes the double helix.
| |||
| |||

Sources of deoxyuridine in DNA include the presence of dUTP because dUMP (the substrate for thymidylate synthase) or dUDP (from ribonucleotide reductase action on UDP) are phosphorylated to the triphosphate. DNA polymerase recognizes these compounds as substrates. Another source is the deamination of deoxycytidine in DNA, promoted by a variety of compounds. If deoxyuridine is on a template strand of the DNA, it will direct the incorporation of an A in the newly made strand of DNA. This will convert a G-C pair to an A-T pair.
Cells have evolved ways of preventing mutations caused by either mechanism. First, cells contain an enzyme, dUTPase,which hydro-lyses dUTP to dUMP. Thus the triphosphate substrate is taken away from DNA polymerase before it can serve as a substrate. Secondly, U residues in DNA from whatever source cause the gap repair system to be activated so that these alterations are removed before they can be replicated. The base-pairing in DNA of dU and dT are identical; each forms two hydrogen bonds with dA on the opposite strand. However, dT is used in DNA; this base contains a methyl group that dU and dC do not. This means that any time dU arises from the loss of the amino group of dC, it can be recognized and removed. If the base 5-methyl C were to be deaminated, that would lead to a T in the DNA. In this case, the repair system couldn't determine which strand of the double helix was incorrect; it would be likely to delete either the altered base or its partner, creating a high probability of mutation at that position. In fact, DNA is often methylated at some sequences—these sites on DNA are often mutational hot spots.
0 comments:
Post a Comment